

Back |
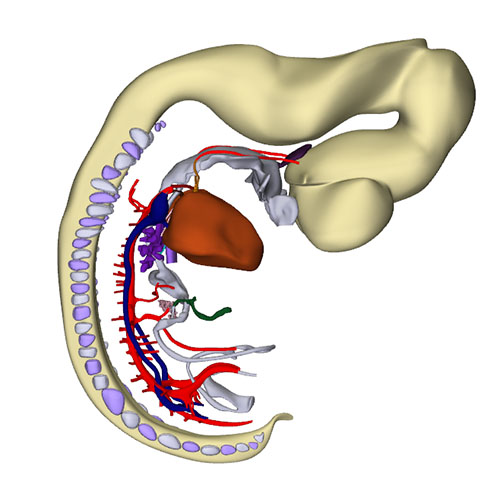
Interactive 3D PDF of Carnegie Stage 17 |
The interactive PDF below has been very kindly provided by Dr. Jill PJM Hikspoors, Prof. Wouter H Lamers,
Department of Anatomy & Embryology, Maastricht University, Maastricht, The Netherlands
Contact: jill.hikspoors@maastrichtuniversity.nl
Publication: 10.1038/s42003-022-03153-x
*Important* Although some web browsers will allow PDFs to be viewed online, the file must be downloaded and saved to a computer to enable the interactive options.
 |
 |
 |
 |
 |
||||
Right-sided view |
Right sagittal view: Closure of the interatrial foramen and appearance of the secondary foramen |
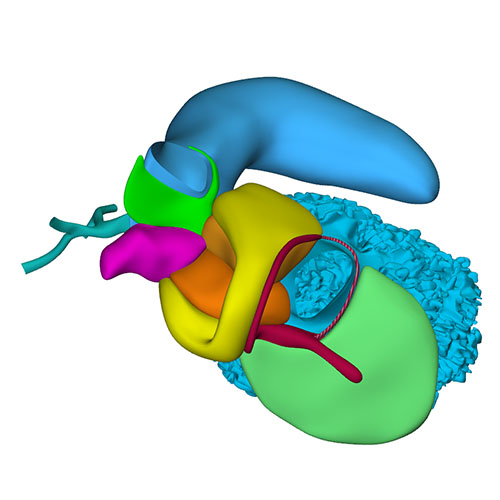
Cranial view: Cardiac conduction system visualizes the expansion of the tricuspid gully rightwards, while subaortic part of the outflow tract remains defined on midline |
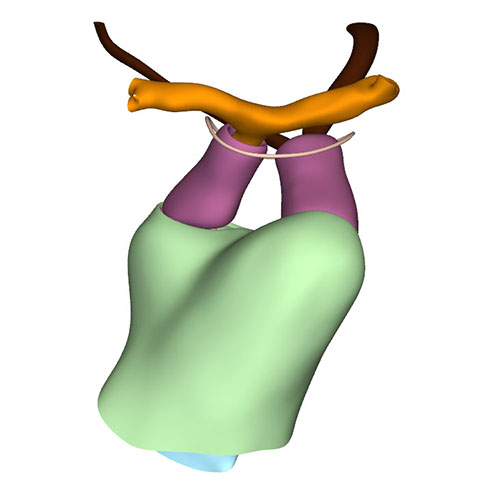
Right ventral view: Extensive changes at arterial pole when compared to CS16 |
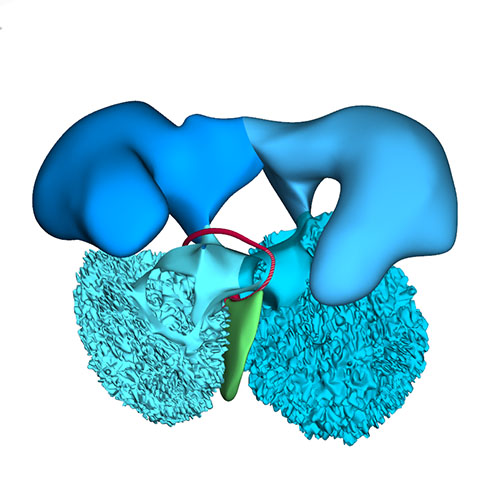
Cranial view: Wedge-shaped ridges visualizing leaflet formation |
||||
 |
 |
 |
 |
|||
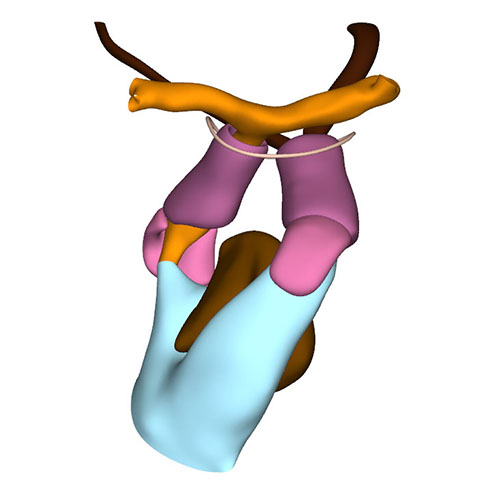
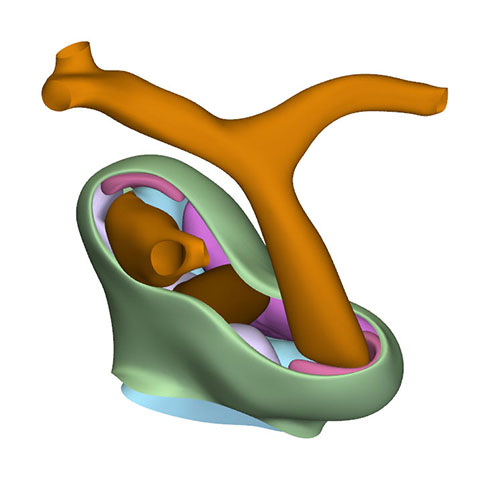
Cranial view: Columns form intrapericardial arterial walls |
Cranial view: Myocardial outflow tract no longer reaches pericardial reflection but retains fish-mouth shape |
Dorsal view: Rotation of the arterial trunks relative to each other and asymmetric growth of the aortic trunk |
Left cranial view: Position of the pulmonary and aortic channels |
At this stage, ~40 days have passed since fertilization. Compared to preceding stages, limited changes were noted in the arrangement of the systemic venous sinus. The superior and inferior atrioventricular cushions, including their atrial extensions, have fused in nearly all embryos at this stage, thus closing the primary atrial foramen. Accordingly, a wide secondary foramen has formed due to breakdown of the dorsal portion of the primary atrial septum in these hearts. The site of fusion of the atrioventricular cushions in the atrium is reinforced on its right side by the dorsal mesenchymal protrusion, or vestibular spine. Myocardium surrounds the stem of the pulmonary vein, which from this stage onwards begins to expand radially, suggesting an increase in blood flow. Small lateral cushions have appeared in the left and right margins of the atrioventricular canal.
The well-developed trabeculations of left and right ventricles form a complex 3-dimensional network, and have developed extensive gap-junctional contacts and myofibers. These properties reflect their faster conduction and stronger contraction. The appearance of a compact left ventricular myocardial wall shows that multiplication of cardiomyocytes at the epicardial side of the ventricular walls now exceeds that in the inner trabecular layer. The compact myocardium does not arise, as is often suggested, by condensation of the trabecular network. While the muscular ventricular septum develops equally from right- and left-ventricular contributions during the phase of trabecular growth, the contribution of cardiomyocytes now becomes proportional to growth in the compact ventricular walls. The peri-tricuspid part of the interventricular foramen, encircling the right atrioventricular orifice, continues to expand in a rightward direction, while its subaortic part remains bordered cranially by the inner curvature and caudally by the crest of the muscular ventricular septum.
Between CS15 and CS20, the volume of the myocardium of the outflow tract hardly increases in both rodent and human embryos. In overall architecture, nonetheless, it undergoes an impressive change in appearance. Its length decreases abruptly to <50% of its original length in the ~2 days separating CS16 from CS17. Much of the effective shortening can be attributed to it changing from a long tubular configuration at CS16 to a shorter figure-of-eight configuration at CS17. The waist of the figure-of-eight corresponds with the developing medial walls of the subpulmonary and subaortic channels. In addition, the proximal portion of the outflow tract itself becomes progressively more wedge-shaped, with its greatest length along the subpulmonary channel and its shortest length along the subaortic channel. The proximal portion of the outflow tract of birds undergoes an almost identical change in appearance at a comparable stage of heart development. The changing shape reflects the ongoing incorporation of the smooth-walled proximal outflow tract into the right ventricle as its infundibulum, and into the left ventricle as its aortic vestibule.
The aortopulmonary septum has fused with the septal and parietal ridges. The fusion of the endocardial ridges mutually now has passed through the middle portion of the outflow tract. As a result, the neural-crest cells of the aortopulmonary septum and the prongs become sequestered inside the ridges as a dense central “whorl” of cells between the anlagen of the arterial valves. The developing aortic and pulmonary roots themselves occupy right-caudal and left-cranial positions, respectively, which represents their definitive position. The distal parts of the septal and parietal ridges undergo a similar change in shape as the corresponding part of the myocardial outflow tract. They now each resemble a heart-shaped structure with wide downstream “ears” separated by a median furrow that presages its separation and allocation to the left and right semilunar leaflets of the aortic and pulmonary roots, respectively. The lateral swellings have also increased markedly in size at this stage. Concomitantly with the distal widening of the ridges and swellings, there is movement in their position relative to the distal orifices of the subaortic and subpulmonary channels. The ridges in the proximal outflow tract have still to fuse.
Since the myocardium of the outflow tract hardly proliferates, and the addition of new cardiomyocytes from the second heart field has ceased, the fishmouth has lost its characteristic appearance. The non-myocardial cells that continue to populate the distal outflow tract, now interpose between the distal margin of the myocardial walls and the pericardial reflection. Consequently, the myocardial boundary moves away from the pericardial reflection.
Concomitant with the marked reduction in length of the myocardial outflow tract, the pulmonary trunk ceases to grow. The ascending aorta, in contrast, continues to increase in length well beyond CS23. The inner layer of the smooth muscular wall of both arterial trunks derives from the neural crest, and the outer layer from the second heart field. However, the wall of the ascending aorta is mainly derived from the neural crest, whereas the wall of the pulmonary trunk originates predominantly in the second heart field. These different contributions may well explain the different growth characteristics of the vessels.
The epicardium that covers the intrapericardial arterial trunks expresses the morphological and molecular features of the nearby pericardium. It consists of a sheet of densely packed cuboidal cells rather than of squamous cells, which characterize the epicardium. It also expresses genes that characterize the pericardium. This “arterial” epicardium seems to form locally, since it spreads across the arterial pole even after the removal of the pro-epicardial body. Collectively, these data indicate that the arterial pole of the heart derives from the ventral wall of the pharynx, with the non-myocardial tissues entering the walls of the distal part of the intrapericardial outflow tract to form its arterial component.
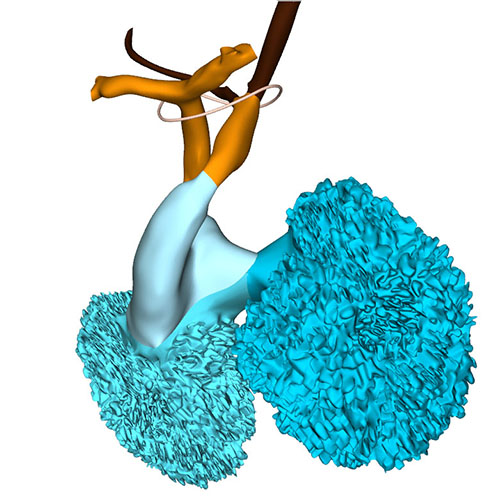
Extensive changes have occurred at the arterial pole. Both dorsal aortas are still present, but the left and right carotid ducts have disappeared. Marked narrowing is now seen in the diameter of the right-sided dorsal aorta between the take-off of the 7th cervical segmental artery and its confluence with the left-sided dorsal aorta, while the lumen of the artery of the right 6th arch has all but obliterated distal to the origin of the right pulmonary artery.
Click an image to download a 3D-PDF. The file must be saved to a computer to enable the interactive options. The 3D-PDFs can be opened on any computer as long as Adobe PDF or equivalent reader is installed.
A 3D-PDF becomes activated by “clicking” with the mouse on the reconstruction.
A toolbar appears at the top of the screen that includes the option “model tree”.
The model tree displays a material list of structures in the upper box, and preset viewing options (cameras) in the lower box. The sequence of items corresponds to that in Supplemental Table 3 of the publication.
The list of visible structures can be modified by marking or unmarking a structure.
To manipulate the reconstruction, press the left mouse button to rotate it, the scroll button to zoom in or out, and the left and right mouse buttons simultaneously to move the embryo across the screen.
A structure can be rendered transparent by selecting that option from the drop-down menu after selecting the structure with the right mouse button.
To inspect a combination of structures, one is advised to build up the composition, beginning with a familiar component, such as a lumen, rather than deleting non-relevant structures one-by-one from a completely reconstructed specimen.
The slicer button in the toolbar allows making cross sections. The plane of section can be adjusted with the offset and tilt options.
The “loop wires” in Supplemental Figures 3-6 of the publication, which are drawn through the center of the endocardial heart tube, emphasize the changing shape of the heart loop during CS10-13.
The side length of the scale cubes is 200 μm.
The preset views correspond to the images shown in Figures 1-10 of the publication.
Note that items that are visible in these views can be altered by marking or unmarking a structure in the model tree.