

Back |
Interactive 3D PDF of Carnegie Stage 9 |
The interactive PDF below has been very kindly provided by Dr. Jill PJM Hikspoors, Prof. Wouter H Lamers,
Department of Anatomy & Embryology, Maastricht University, Maastricht, The Netherlands
Contact: jill.hikspoors@maastrichtuniversity.nl
Publication: 10.1038/s42003-022-03153-x
*Important* Although some web browsers will allow PDFs to be viewed online, the file must be downloaded and saved to a computer to enable the interactive options.
 |
 |
 |
 |
|||
Right-sided view |
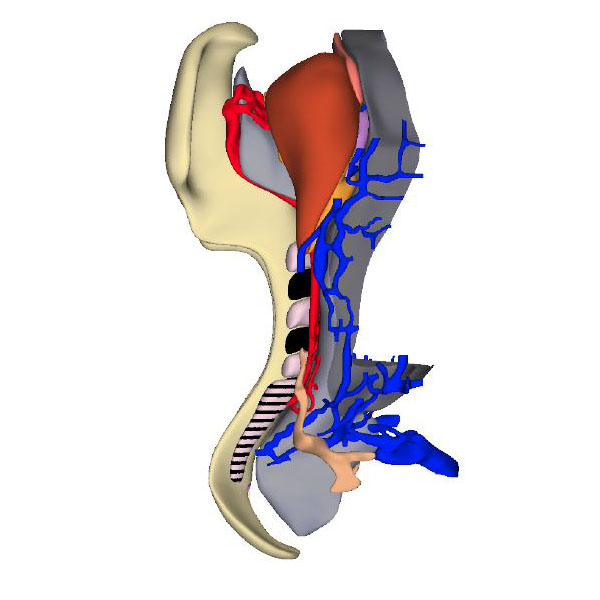
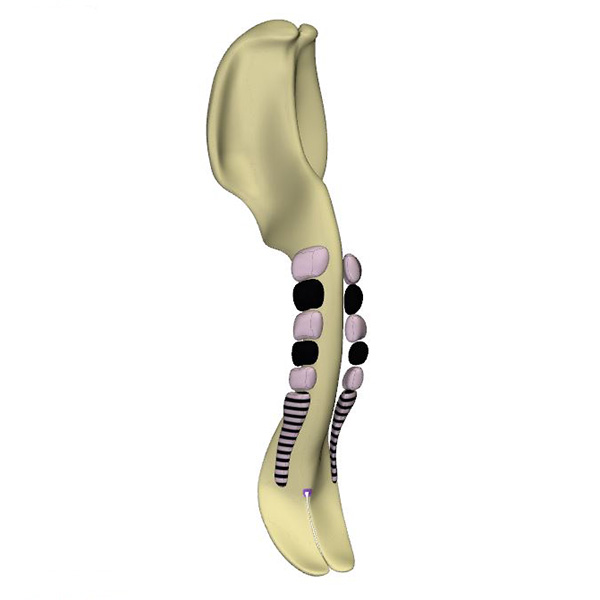
Right ventral view: Neural plate with Hensen's node and flanked by 5 somites |
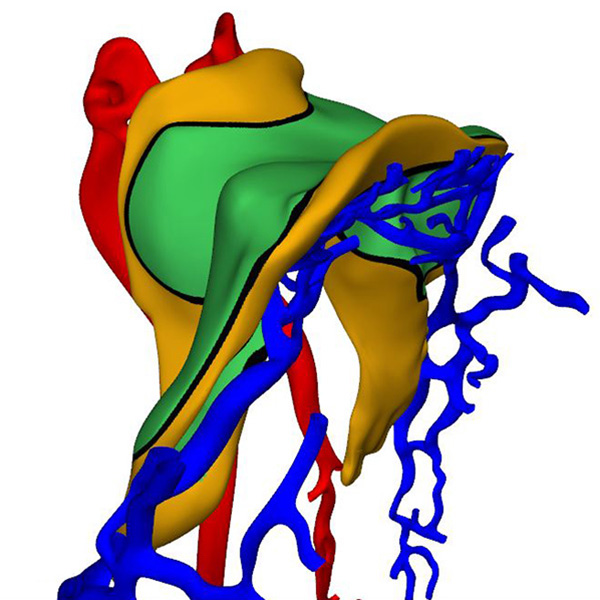
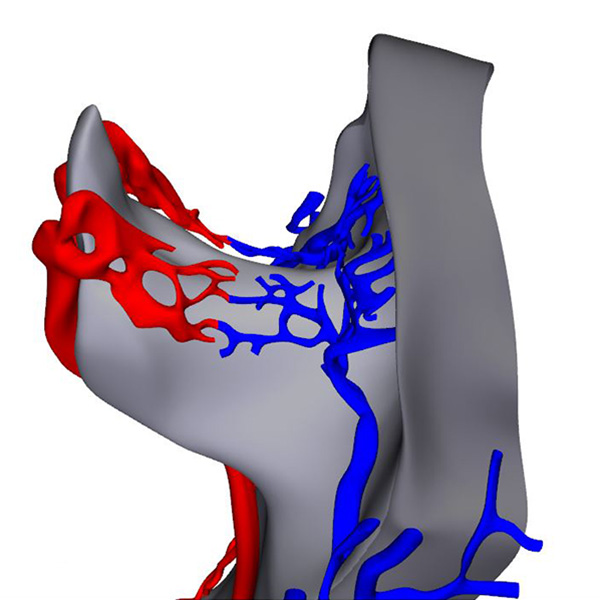
Right ventral view: Horseshoe-shaped visceral pericardial wall |
Right ventral view: Cardiac primordia are bilaterally symmetrical |
The heart becomes morphologically identifiable when ~26 days have passed since fertilization. This embryo has developed a neural plate that is flanked by 5 somites and somitomeres. The endoderm, shown in grey, is forming the pharynx. It is continuous at its periphery with the yolk sac, shown in darker grey. The horseshoe-shaped pericardial cavity covers the endoderm in front and laterally, where it becomes gradually narrower to end adjacent to the first somite. Gastrulation begins during CS8 (~23 days of development). By CS9, Hensen’s node, which localizes gastrulation, is found at the caudal end of the columns of somitomeres.
The heart is located at the cranial margin of the embryo. At CS8, midway through gastrulation, bilateral heart fields, also known as cardiogenic plates, form craniolaterally in the embryonic mesoderm. The cranial part of these morphologically still indistinct precursors forms the “first heart field” and differentiates to form cardiomyocytes during or shortly after the formation of the first somite. The remaining, more caudally located and still proliferating precursors, which form the “second heart field”, start to differentiate at the transition of CS9 into CS10. Cardiomyocyte contractions arise in the lateral regions of the cardiac crescents of mouse embryos as soon as sarcomeric assembly and Ca2+ transients can be demonstrated. Visible contractions develop in mouse embryos when 3 somites have formed, while the first cardiac contractions in human embryos can be visualized with ultrasound at days 28 after conception. The first heart field can be visualized in mice by the expression of transcription factors Tbx5 and Hcn4, and the second heart field by the expression of the transcription factor Isl1. Fate-mapping studies in mice have shown that the first heart field contributes virtually all cardiomyocytes of the embryonic left ventricle, three-quarters of those in the atrioventricular canal, and half of those making up the atriums and right ventricle. The more slowly evolving second heart field, on the other hand, contributes virtually all cardiomyocytes of the outflow tract, half of the atria and right ventricles, and one quarter of the atrioventricular canal. The systemic venous sinus is a separate part of the second heart field that does not express the early cardiogenic transcription factor Nkx2-5, but does express Tbx18. Based on molecular mouse data, we infer that the center of the heart develops first, and that the upstream venous and downstream arterial components are added successively.
The 3D-PDF and other reconstructions of embryos with ~5 somites, show that the early heart is, like that in mouse embryos, bilaterally symmetrical. It consists of two tiny vascular networks that course in front of the foregut, with only few vessels connecting them across the midline. Both vascular networks are surrounded by paired, but partially merged swellings of acellular cardiac jelly. The jelly is enclosed, in turn, within an unpaired, bilaterally symmetrical pericardial cavity with a myogenic visceral wall. The distribution of the jelly, which is produced by endoderm and the visceral pericardial wall, reflects the location of the boundaries of the differentiating left ventricular myocardium. The initially non-luminal endocardial tubes of the cardiac vascular network gradually canalize, but at first contain only few erythrocytes. At the venous pole, the heart tubes are continuous with an extensive venous plexus on the periphery of the endoderm. At the arterial pole, near the buccopharyngeal membrane, the heart tubes pass the pharynx laterally to join the paired dorsal aortas. In front of the heart, the primordium of the transverse septum forms as a shelf of thick mesoderm between the endoderm and the pericardial cavity.
Click an image to download a 3D-PDF. The file must be saved to a computer to enable the interactive options. The 3D-PDFs can be opened on any computer as long as Adobe PDF or equivalent reader is installed.
A 3D-PDF becomes activated by “clicking” with the mouse on the reconstruction.
A toolbar appears at the top of the screen that includes the option “model tree”.
The model tree displays a material list of structures in the upper box, and preset viewing options (cameras) in the lower box. The sequence of items corresponds to that in Supplemental Table 3 of the publication.
The list of visible structures can be modified by marking or unmarking a structure.
To manipulate the reconstruction, press the left mouse button to rotate it, the scroll button to zoom in or out, and the left and right mouse buttons simultaneously to move the embryo across the screen.
A structure can be rendered transparent by selecting that option from the drop-down menu after selecting the structure with the right mouse button.
To inspect a combination of structures, one is advised to build up the composition, beginning with a familiar component, such as a lumen, rather than deleting non-relevant structures one-by-one from a completely reconstructed specimen.
The slicer button in the toolbar allows making cross sections. The plane of section can be adjusted with the offset and tilt options.
The “loop wires” in Supplemental Figures 3-6 of the publication, which are drawn through the center of the endocardial heart tube, emphasize the changing shape of the heart loop during CS10-13.
The side length of the scale cubes is 200 μm.
The preset views correspond to the images shown in Figures 1-10 of the publication.
Note that items that are visible in these views can be altered by marking or unmarking a structure in the model tree.